




The nervous system of the common laboratory fly, Drosophila melanogaster, contains around 100,000 neurons, the same number as a lobster. This number compares to 75 million in the mouse and 300 million in the octopus. A human brain contains around 86 billion neurons. Despite these very different numbers, the nervous systems of these animals control many of the same behaviors—from basic reflexes to more complicated behaviors like finding food and courting mates. The ability of neurons to communicate with each other as well as with other types of cells underlies all of these behaviors.
Most neurons share the same cellular components. But neurons are also highly specialized—different types of neurons have different sizes and shapes that relate to their functional roles.
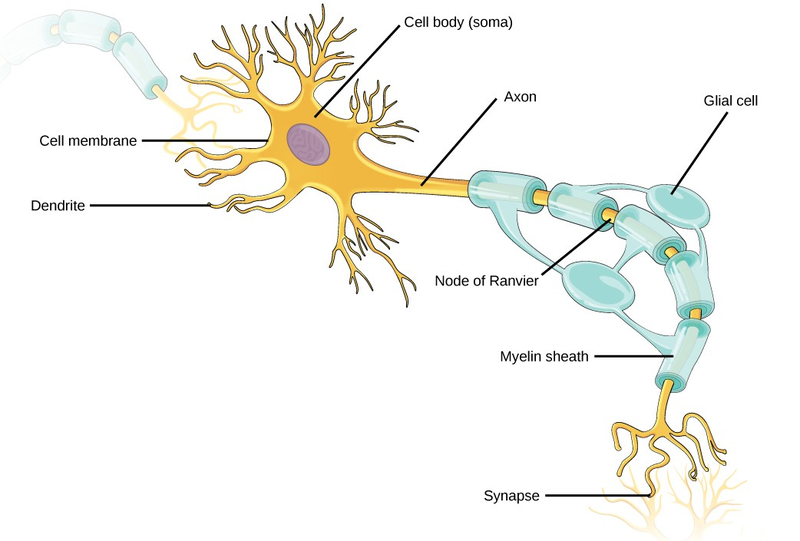
Like other cells, each neuron has a cell body (or soma) that contains a nucleus, smooth and rough endoplasmic reticulum, Golgi apparatus, mitochondria, and other cellular components. Neurons also contain unique structures for receiving and sending the electrical signals that make communication between neurons possible (Figure 16.19). Dendrites are tree-like structures that extend away from the cell body to receive messages from other neurons at specialized junctions called synapses. Although some neurons do not have any dendrites, most have one or many dendrites.
The bilayer lipid membrane that surrounds a neuron is impermeable to ions. To enter or exit the neuron, ions must pass through ion channels that span the membrane. Some ion channels need to be activated to open and allow ions to pass into or out of the cell. These ion channels are sensitive to the environment and can change their shape accordingly. Ion channels that change their structure in response to voltage changes are called voltage-gated ion channels. The difference in total charge between the inside and outside of the cell is called the membrane potential.
A neuron at rest is negatively charged: the inside of a cell is approximately 70 millivolts more negative than the outside (–70 mV). This voltage is called the resting membrane potential; it is caused by differences in the concentrations of ions inside and outside the cell and the selective permeability created by ion channels. Sodium-potassium pumps in the membrane produce the different ion concentrations inside and outside of the cell by bringing in two K+ ions and removing three Na+ ions. The actions of this pump are costly: one molecule of ATP is used up for each turn. Up to 50 percent of a neuron’s ATP is used in maintaining its membrane resting potential. Potassium ions (K+), which are higher inside the cell, move fairly freely out of the neuron through potassium channels; this loss of positive charge produces a net negative charge inside the cell. Sodium ions (Na+), which are low inside, have a driving force to enter but move less freely. Their channels are voltage dependent and will open when a slight change in the membrane potential triggers them.
A neuron can receive input from other neurons and, if this input is strong enough, send the signal to downstream neurons. Transmission of a signal between neurons is generally carried by a chemical, called a neurotransmitter, which diffuses from the axon of one neuron to the dendrite of a second neuron. When neurotransmitter molecules bind to receptors located on a neuron’s dendrites, the neurotransmitter opens ion channels in the dendrite’s plasma membrane. This opening allows sodium ions to enter the neuron and results in depolarization of the membrane—a decrease in the voltage across the neuron membrane. Once a signal is received by the dendrite, it then travels passively to the cell body. A large enough signal from neurotransmitters will reach the axon. If it is strong enough (that is, if the threshold of excitation, a depolarization to around –60mV is reached), then depolarization creates a positive feedback loop: as more Na+ ions enter the cell, the axon becomes further depolarized, opening even more sodium channels at further distances from the cell body. This will cause voltage dependent Na+ channels further down the axon to open and more positive ions to enter the cell. In the axon, this “signal” will become a self- propagating brief reversal of the resting membrane potential called an action potential.
An action potential is an all-or-nothing event; it either happens or it does not. The threshold of excitation must be reached for the neuron to “fire” an action potential. As sodium ions rush into the cell, depolarization actually reverses the charge across the membrane form -70mv to +30mV. This change in the membrane potential causes voltage-gated K+ channels to open, and K+ begins to leave the cell, repolarizing it. At the same time, Na+ channels inactivate so no more Na+ enters the cell. K+ ions continue to leave the cell and the membrane potential returns to the resting potential. At the resting potential, the K+ channels close and Na+ channels reset. The depolarization of the membrane proceeds in a wave down the length of the axon. It travels in only one direction because the sodium channels have been inactivated and unavailable until the membrane potential is near the resting potential again; at this point they are reset to closed and can be opened again.
An axon is a tube-like structure that propagates the signal from the cell body to specialized endings called axon terminals. These terminals in turn then synapse with other neurons, muscle, or target organs. When the action potential reaches the axon terminal, this causes the release of neurotransmitter onto the dendrite of another neuron. Neurotransmitters released at axon terminals allow signals to be communicated to these other cells, and the process begins again. Neurons usually have one or two axons, but some neurons do not contain any axons.
Some axons are covered with a special structure called a myelinsheath, which acts as an insulator to keep the electrical signal from dissipating as it travels down the axon. This insulation is important, as the axon from a human motor neuron can be as long as a meter (3.2 ft)—from the base of the spine to the toes. The myelin sheath is produced by glial cells. Along the axon there are periodic gaps in the myelin sheath. These gaps are called nodes of Ranvier and are sites where the signal is “recharged” as it travels along the axon.
It is important to note that a single neuron does not act alone—neuronal communication depends on the connections that neurons make with one another (as well as with other cells, like muscle cells). Dendrites from a single neuron may receive synaptic contact from many other neurons. For example, dendrites from a Purkinje cell in the cerebellum are thought to receive contact from as many as 200,000 other neurons.
Biology IN ACTION
Neurogenesis
At one time, scientists believed that people were born with all the neurons they would ever have. Research performed during the last few decades indicates that neurogenesis, the birth of new neurons, continues into adulthood. Neurogenesis was first discovered in songbirds that produce new neurons while learning songs. For mammals, new neurons also play an important role in learning: about 1,000 new neurons develop in the hippocampus (a brain structure involved in learning and memory) each day. While most of the new neurons will die, researchers found that an increase in the number of surviving new neurons in the hippocampus correlated with how well rats learned a new task. Interestingly, both exercise and some antidepressant medications also promote neurogenesis in the hippocampus. Stress has the opposite effect. While neurogenesis is quite limited compared to regeneration in other tissues, research in this area may lead to new treatments for disorders such as Alzheimer’s, stroke, and epilepsy.
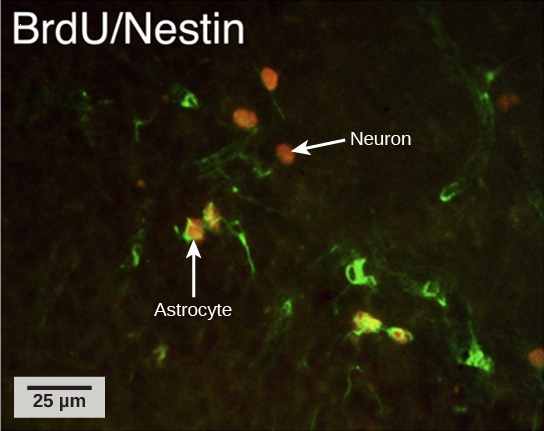
How do scientists identify new neurons? A researcher can inject a compound called bromodeoxyuridine (BrdU) into the brain of an animal. While all cells will be exposed to BrdU, BrdU will only
be incorporated into the DNA of newly generated cells that are in S phase. A technique called immunohistochemistry can be used to attach a fluorescent label to the incorporated BrdU, and a
researcher can use fluorescent microscopy to visualize the presence of BrdU, and thus new neurons, in brain tissue (Figure 16.20).
Visit this link interactive lab (http://openstaxcollege.org/l/neurogenesis2)to see more information about neurogenesis, including an interactive laboratory simulation and a video that explains how BrdU labels new cells.
While glial cells are often thought of as the supporting cast of the nervous system, the number of glial cells in the brain actually outnumbers the number of neurons by a factor of 10. Neurons would be unable to function without the vital roles that are fulfilled by these glial cells. Glia guide developing neurons to their destinations, buffer ions and chemicals that would otherwise harm neurons, and provide myelin sheaths around axons. When glia do not function properly, the result can be disastrous—most brain tumors are caused by mutations in glia.
- 3785 reads
