




Recall that a gene for a particular character may have several variants, or alleles, that code for different traits associated with that character. For example, in the ABO blood type system in humans, three alleles determine the particular blood-type protein on the surface of red blood cells. Each individual in a population of diploid organisms can only carry two alleles for a particular gene, but more than two may be present in the individuals that make up the population. Mendel followed alleles as they were inherited from parent to offspring. In the early twentieth century, biologists began to study what happens to all the alleles in a population in a field of study known as population genetics.
Until now, we have defined evolution as a change in the characteristics of a population of organisms, but behind that phenotypic change is genetic change. In population genetic terms, evolution is defined as a change in the frequency of an allele in a population. Using the ABO system as an example, the frequency of one of the alleles, IA, is the number of copies of that allele divided by all the copies of the ABO gene in the population. For example, a study in Jordan found a frequency of IA to be 26.1 percent. 1 The IB,I0 alleles made up 13.4 percent and 60.5 percent of the alleles respectively, and all of the frequencies add up to 100 percent. A change in this frequency over time would constitute evolution in the population.
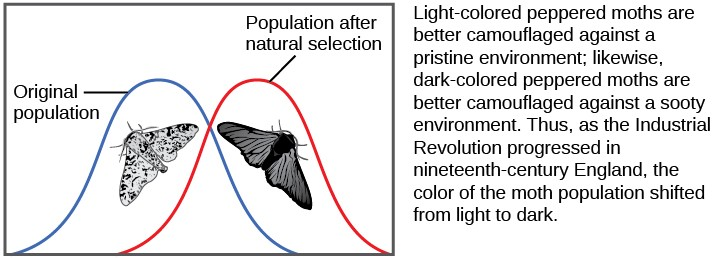
There are several ways the allele frequencies of a population can change. One of those ways is natural selection. If a given allele confers a phenotype that allows an individual to have more offspring that survive and reproduce, that allele, by virtue of being inherited by those offspring, will be in greater frequency in the next generation. Since allele frequencies always add up to 100 percent, an increase in the frequency of one allele always means a corresponding decrease in one or more of the other alleles. Highly beneficial alleles may, over a very few generations, become “fixed” in this way, meaning that every individual of the population will carry the allele. Similarly, detrimental alleles may be swiftly eliminated from the gene pool, the sum of all the alleles in a population. Part of the study of population genetics is tracking how selective forces change the allele frequencies in a population over time, which can give scientists clues regarding the selective forces that may be operating on a given population. The studies of changes in wing coloration in the peppered moth from mottled white to dark in response to soot-covered tree trunks and then back to mottled white when factories stopped producing so much soot is a classic example of studying evolution in natural populations (Figure 11.6).
In the early twentieth century, English mathematician Godfrey Hardy and German physician Wilhelm Weinberg independently provided an explanation for a somewhat counterintuitive concept. Hardy’s original explanation was in response to a misunderstanding as to why a “dominant” allele, one that masks a recessive allele, should not increase in frequency in a population until it eliminated all the other alleles. The question resulted from a common confusion about what “dominant” means, but it forced Hardy, who was not even a biologist, to point out that if there are no factors that affect an allele frequency those frequencies will remain constant from one generation to the next. This principle is now known as the Hardy-Weinberg equilibrium. The theory states that a population’s allele and genotype frequencies are inherently stable—unless some kind of evolutionary force is acting on the population, the population would carry the same alleles in the same proportions generation after generation. Individuals would, as a whole, look essentially the same and this would be unrelated to whether the alleles were dominant or recessive. The four most important evolutionary forces, which will disrupt the equilibrium, are natural selection, mutation, genetic drift, and migration into or out of a population. A fifth factor, nonrandom mating, will also disrupt the Hardy-Weinberg equilibrium but only by shifting genotype frequencies, not allele frequencies. In nonrandom mating, individuals are more likely to mate with like individuals (or unlike individuals) rather than at random. Since nonrandom mating does not change allele frequencies, it does not cause evolution directly. Natural selection has been described. Mutation creates one allele out of another one and changes an allele’s frequency by a small, but continuous amount each generation. Each allele is generated by a low, constant mutation rate that will slowly increase the allele’s frequency in a population if no other forces act on the allele. If natural selection acts against the allele, it will be removed from the population at a low rate leading to a frequency that results from a balance between selection and mutation. This is one reason that genetic diseases remain in the human population at very low frequencies. If the allele is favored by selection, it will increase in frequency. Genetic drift causes random changes in allele frequencies when populations are small. Genetic drift can often be important in evolution, as discussed in the next section. Finally, if two populations of a species have different allele frequencies, migration of individuals between them will cause frequency changes in both populations. As it happens, there is no population in which one or more of these processes are not operating, so populations are always evolving, and the Hardy-Weinberg equilibrium will never be exactly observed. However, the Hardy-Weinberg principle gives scientists a baseline expectation for allele frequencies in a non-evolving population to which they can compare evolving populations and thereby infer what evolutionary forces might be at play. The population is evolving if the frequencies of alleles or genotypes deviate from the value expected from the Hardy-Weinberg principle.
Darwin identified a special case of natural selection that he called sexual selection. Sexual selection affects an individual’s ability to mate and thus produce offspring, and it leads to the evolution of dramatic traits that often appear maladaptive in terms of survival but persist because they give their owners greater reproductive success. Sexual selection occurs in two ways: through male–male competition for mates and through female selection of mates. Male–male competition takes the form of conflicts between males, which are often ritualized, but may also pose significant threats to a male’s survival. Sometimes the competition is for territory, with females more likely to mate with males with higher quality territories. Female choice occurs when females choose a male based on a particular trait, such as feather colors, the performance of a mating dance, or the building of an elaborate structure. In some cases male–male competition and female choice combine in the mating process. In each of these cases, the traits selected for, such as fighting ability or feather color and length, become enhanced in the males. In general, it is thought that sexual selection can proceed to a point at which natural selection against a character’s further enhancement prevents its further evolution because it negatively impacts the male’s ability to survive. For example, colorful feathers or an elaborate display make the male more obvious to predators.
- 2725 reads
