




Viruses were first discovered after the development of a porcelain filter, called the Chamberland- Pasteur filter, which could remove all bacteria visible under the microscope from any liquid sample. In 1886, Adolph Meyer demonstrated that a disease of tobacco plants, tobacco mosaic disease, could be transferred from a diseased plant to a healthy one through liquid plant extracts. In 1892, Dmitri Ivanowski showed that this disease could be transmitted in this way even after the Chamberland-Pasteur filter had removed all viable bacteria from the extract. Still, it was many years before it was proven that these “filterable” infectious agents were not simply very small bacteria but were a new type of tiny, disease-causing particle.
Virions, single virus particles, are very small, about 20–250 nanometers (1 nanometer = 1/1,000,000 mm). These individual virus particles are the infectious form of a virus outside the host cell. Unlike bacteria (which are about 100 times larger), we cannot see viruses with a light microscope, with the exception of some large virions of the poxvirus family (Figure 17.3).
It was not until the development of the electron microscope in the 1940s that scientists got their first good view of the structure of the tobacco mosaic virus (Figure 17.2) and others. The surface structure of virions can be observed by both scanning and transmission electron microscopy, whereas the internal structures of the virus can only be observed in images from a transmission electron microscope (Figure 17.4).
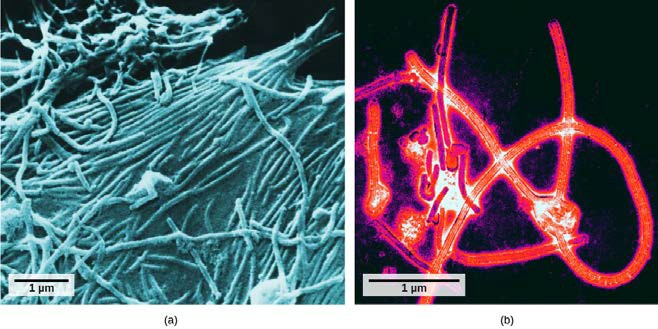
The use of this technology has allowed for the discovery of many viruses of all types of living organisms. They were initially grouped by shared morphology, meaning their size, shape, and distinguishing structures. Later, groups of viruses were classified by the type of nucleic acid they contained, DNA or RNA, and whether their nucleic acid was single- or double-stranded. More recently, molecular analysis of viral replication cycles has further refined their classification.
A virion consists of a nucleic-acid core, an outer protein coating, and sometimes an outer envelope made of protein and phospholipid membranes derived from the host cell. The most visible difference between members of viral families is their morphology, which is quite diverse. An interesting feature of viral complexity is that the complexity of the host does not correlate to the complexity of the virion. Some of the most complex virion structures are observed in bacteriophages, viruses that infect the simplest living organisms, bacteria.
Viruses come in many shapes and sizes, but these are consistent and distinct for each viral family (Figure 17.5). All virions have a nucleic-acid genome covered by a protective layer of protein, called a capsid. The capsid is made of protein subunits called capsomeres. Some viral capsids are simple polyhedral “spheres,” whereas others are quite complex in structure. The outer structure surrounding the capsid of some viruses is called the viralenvelope. All viruses use some sort of glycoprotein to attach to their host cells at molecules on the cell called viral receptors. The virus exploits these cell-surface molecules, which the cell uses for some other purpose, as a way to recognize and infect specific cell types. For example, the measles virus uses a cell-surface glycoprotein in humans that normally functions in immune reactions and possibly in the sperm-egg interaction at fertilization. Attachment is a requirement for viruses to later penetrate the cell membrane, inject the viral genome, and complete their replication inside the cell.
The T4 bacteriophage, which infects the E.colibacterium, is among the most complex virion known; T4 has a protein tail structure that the virus uses to attach to the host cell and a head structure that houses its DNA.
Adenovirus, a nonenveloped animal virus that causes respiratory illnesses in humans, uses protein spikes protruding from its capsomeres to attach to the host cell. Nonenveloped viruses also include those that cause polio (poliovirus), plantar warts (papillomavirus), and hepatitis A (hepatitis A virus). Nonenveloped viruses tend to be more robust and more likely to survive under harsh conditions, such as the gut.
Enveloped virions like HIV (human immunodeficiency virus), the causative agent in AIDS (acquired immune deficiency syndrome), consist of nucleic acid (RNA in the case of HIV) and capsid proteins surrounded by a phospholipid bilayer envelope and its associated proteins (Figure 17.5). Chicken pox, influenza, and mumps are examples of diseases caused by viruses with envelopes. Because of the fragility of the envelope, nonenveloped viruses are more resistant to changes in temperature, pH, and some disinfectants than enveloped viruses.
Overall, the shape of the virion and the presence or absence of an envelope tells us little about what diseases the viruses may cause or what species they might infect, but is still a useful means to begin viral classification.
Art CONNECTION
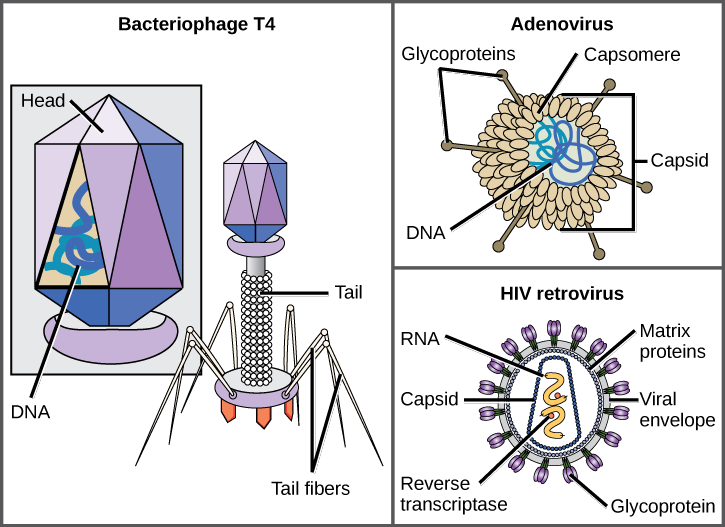
Which of the following statements about virus structure is true?
a. All viruses are encased in a viral membrane.
b. The capsomere is made up of small protein subunits called capsids.
c. DNA is the genetic material in all viruses.
d. Glycoproteins help the virus attach to the host cell.
Unlike all living organisms that use DNA as their genetic material, viruses may use either DNA or RNA as theirs. The virus core contains the genome or total genetic content of the virus. Viral genomes tend to be small compared to bacteria or eukaryotes, containing only those genes that code for proteins the virus cannot get from the host cell. This genetic material may be single-stranded or double-stranded. It may also be linear or circular. While most viruses contain a single segment of nucleic acid, others have genomes that consist of several segments.
DNA viruses have a DNA core. The viral DNA directs the host cell’s replication proteins to synthesize new copies of the viral genome and to transcribe and translate that genome into viral proteins. DNA viruses cause human diseases such as chickenpox, hepatitis B, and some venereal diseases like herpes and genital warts.
RNA viruses contain only RNA in their cores. To replicate their genomes in the host cell, the genomes of RNA viruses encode enzymes not found in host cells. RNA polymerase enzymes are not as stable as DNA polymerases and often make mistakes during transcription. For this reason, mutations, changes in the nucleotide sequence, in RNA viruses occur more frequently than in DNA viruses. This leads to more rapid evolution and change in RNA viruses. For example, the fact that influenza is an RNA virus is one reason a new flu vaccine is needed every year. Human diseases caused by RNA viruses include hepatitis C, measles, and rabies.
Viruses can be seen as obligate intracellular parasites. The virus must attach to a living cell, be taken inside, manufacture its proteins and copy its genome, and find a way to escape the cell so the virus can infect other cells and ultimately other individuals. Viruses can infect only certain species of hosts and only certain cells within that host. The molecular basis for this specificity is that a particular surface molecule, known as the viral receptor, must be found on the host cell surface for the virus to attach. Also, metabolic differences seen in different cell types based on differential gene expression are a likely factor in which cells a virus may use to replicate. The cell must be making the substances the virus needs, such as enzymes the virus genome itself does not have genes for, or the virus will not be able to replicate using that cell.
- 6620 reads
